Background: The pathogenesis of acute kidney injury (AKI) in sepsis involves inflammatory response. Autophagy has been shown to regulate inflammatory response, but the role of autophagy-related genes (ARGs) in the regulation of inflammation in septic AKI requires further investigation. Methods: Initially, the dataset GSE57065 was downloaded and utilized in the R language to investigate differentially expressed autophagy-related genes (DEARGs) associated with septic shock. Subsequently, DEARGs enrichment analysis and protein-protein interactions (PPIs) were conducted. Hub genes were identified through PPIs, and their diagnostic value was evaluated using ROC analyses with the external dataset GSE65682. Additionally, the septic AKI animal model was established to validate hub genes through qRT-PCR. Finally, immune cell infiltration in the septic AKI and control group was analyzed. Furthermore, we examined the correlation between immune cell infiltration and the validated hub genes, and predicted the miRNA-mRNA network. Results: In the GSE57065 dataset, we have identified 22 differentially expressed genes (DEGs) primarily involved in autophagy. The receiver operating characteristic (ROC) analysis suggested that the top ten hub genes may have diagnostic value. In our animal experiment, qRT-PCR results demonstrated elevated expressions of TP53, MYC, FOXO1, CXCR4, and BCL-2, while PARP1 and PTEN exhibited reduced expressions in septic AKI. The CIBERSORT analysis revealed immune infiltration, and the validated hub genes were associated with immune cell infiltration in septic AKI. Lastly, we have discovered potential regulatory miRNAs through the miRNA-mRNA network. Conclusions: The potential diagnostic and therapeutic targets, namely TP53, MYC, FOXO1, CXCR4, BCL-2, PARP1, and PTEN, have been identified as potentially influential factors in the infiltration of immune cells in acute kidney injury associated with sepsis.
| Published in | Biomedical Sciences (Volume 11, Issue 2) |
| DOI | 10.11648/j.bs.20251102.11 |
| Page(s) | 24-35 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Autophagy, Immune Infiltration, Inflammation, Sepsis, Acute Kidney Injury, Bioinformatics Analysis
AKI | Acute Kidney Injury |
ARGs | Utophagy-related Genes |
DEARGs | Differentially Expressed Autophagy-related Genes |
PPIs | Protein-Protein Interactions |
DEGs | Differentially Expressed Genes |
ROC | Receiver Operating Characteristic |
PCA | Principal Component Analysis |
GO | Gene Ontology |
KEGG | Kyoto Encyclopedia of Genes and Genomes |
MCC | Multiscale Curvature Classification |
BP | Biological Process |
CC | Cell Components |
MF | Molecular Function |
BUN | Blood Urea Nitrogen |
| [1] | Singer M, Deutschman CS, Seymour CW, Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche JD, Coopersmith CM, et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). Jama, 2016, 315(8): 801-810. |
| [2] | Delano MJ, Ward PA. The immune system's role in sepsis progression, resolution, and long-term outcome. Immunological reviews, 2016, 274(1): 330-353. |
| [3] | Lee J, Levy MM. Treatment of Patients with Severe Sepsis and Septic Shock: Current Evidence-Based Practices. Rhode Island medical journal (2013), 2019, 102(10): 18-21. |
| [4] | Schmid D, Münz C. Innate and adaptive immunity through autophagy. Immunity, 2007, 27(1): 11-21. |
| [5] | Ho J, Yu J, Wong SH, Zhang L, Liu X, Wong WT, Leung CC, Choi G, Wang MH, Gin T, et al. Autophagy in sepsis: Degradation into exhaustion? Autophagy, 2016, 12(7): 1073-1082. |
| [6] | Gong FC, Ji R, Wang YM, Yang ZT, Chen Y, Mao EQ, Chen EZ. Identification of Potential Biomarkers and Immune Features of Sepsis Using Bioinformatics Analysis. Mediators of inflammation, 2020, 2020: 3432587. |
| [7] | Park SY, Shrestha S, Youn YJ, Kim JK, Kim SY, Kim HJ, Park SH, Ahn WG, Kim S, Lee MG, et al. Autophagy Primes Neutrophils for Neutrophil Extracellular Trap Formation during Sepsis. American journal of respiratory and critical care medicine, 2017, 196(5): 577-589. |
| [8] | Ying L, Zhao GJ, Wu Y, Ke HL, Hong GL, Zhang H, Dong N, Wu Y, Yao YM, Lu ZQ. Mitofusin 2 Promotes Apoptosis of CD4(+) T Cells by Inhibiting Autophagy in Sepsis. Mediators of inflammation, 2017, 2017: 4926205. |
| [9] | Ji Q, Sun Z, Yang Z, Zhang W, Ren Y, Chen W, Yao M, Nie S. Protective effect of ginsenoside Rg1 on LPS-induced apoptosis of lung epithelial cells. Molecular immunology, 2021, 136: 168-174. |
| [10] | Leventhal JS, Ni J, Osmond M, Lee K, Gusella GL, Salem F, Ross MJ. Autophagy Limits Endotoxemic Acute Kidney Injury and Alters Renal Tubular Epithelial Cell Cytokine Expression. PloS one, 2016, 11(3): e0150001. |
| [11] | Wang S, Xia P, Huang G, Zhu P, Liu J, Ye B, Du Y, Fan Z. FoxO1-mediated autophagy is required for NK cell development and innate immunity. Nature communications, 2016, 7: 11023. |
| [12] | Luo J, Wang F, Sun F, Yue T, Zhou Q, Yang C, Rong S, Yang P, Xiong F, Yu Q, et al. Targeted Inhibition of FTO Demethylase Protects Mice Against LPS-Induced Septic Shock by Suppressing NLRP3 Inflammasome. Frontiers in immunology, 2021, 12: 663295. |
| [13] | Cano-Ramos E, Lavin B, Pello OM. Inhibition of MYC in macrophages: tumor vs inflammation-related diseases. Oncoimmunology, 2014, 3(10): e956013. |
| [14] | Zhang Y, Huang T, Jiang L, Gao J, Yu D, Ge Y, Lin S. MCP-induced protein 1 attenuates sepsis-induced acute lung injury by modulating macrophage polarization via the JNK/c-Myc pathway. International immunopharmacology, 2019, 75: 105741. |
| [15] | Sisti F, Wang S, Brandt SL, Glosson-Byers N, Mayo LD, Son YM, Sturgeon S, Filgueiras L, Jancar S, Wong H, et al. Nuclear PTEN enhances the maturation of a microRNA regulon to limit MyD88-dependent susceptibility to sepsis. Science signaling, 2018, 11(528). |
| [16] | Zhang Z, Chen Z, Liu R, Liang Q, Peng Z, Yin S, Tang J, Gong T, Liu Y. Bcl-2 Proteins Regulate Mitophagy in Lipopolysaccharide-Induced Acute Lung Injury via PINK1/Parkin Signaling Pathway. Oxidative medicine and cellular longevity, 2020, 2020: 6579696. |
| [17] | Xu X, Wang H, Liu S, Xing C, Liu Y, Aodengqimuge, Zhou W, Yuan X, Ma Y, Hu M, et al. TP53-dependent autophagy links the ATR-CHEK1 axis activation to proinflammatory VEGFA production in human bronchial epithelial cells exposed to fine particulate matter (PM2.5). Autophagy, 2016, 12(10): 1832-1848. |
| [18] | Ngamsri KC, Jans C, Putri RA, Schindler K, Gamper-Tsigaras J, Eggstein C, Köhler D, Konrad FM. Inhibition of CXCR4 and CXCR7 Is Protective in Acute Peritoneal Inflammation. Frontiers in immunology, 2020, 11: 407. |
| [19] | Wang W, Zhang Y, Liu W, Zhang X, Xiao H, Zhao M, Luo B. CXCR4 induces cell autophagy and maintains EBV latent infection in EBVaGC. Theranostics, 2020, 10(25): 11549-11561. |
| [20] | Soriano FG, Liaudet L, Szabó E, Virág L, Mabley JG, Pacher P, Szabó C. Resistance to acute septic peritonitis in poly (ADP-ribose) polymerase-1-deficient mice. Shock (Augusta, Ga), 2002, 17(4): 286-292. |
| [21] | Singh MP, Chauhan AK, Kang SC. Morin hydrate ameliorates cisplatin-induced ER stress, inflammation and autophagy in HEK-293 cells and mice kidney via PARP-1 regulation. International immunopharmacology, 2018, 56: 156-167. |
| [22] | Kimura T, Jia J, Kumar S, et al. Dedicated SNAREs and specialized TRIM cargo receptors mediate secretory autophagy [J]. The EMBO journal, 2017, 36(1): 42-60. |
| [23] | Qiu P, Liu Y, Zhang J. Review: the Role and Mechanisms of Macrophage Autophagy in Sepsis [J]. Inflammation, 2019, 42(1): 6-19. |
| [24] | Deng Z, Sun M, Wu J, et al. SIRT1 attenuates sepsis-induced acute kidney injury via Beclin1 deacetylation-mediated autophagy activation [J]. Cell death & disease, 2021, 12(2): 217. |
| [25] | Miao S, Lv C, Liu Y, et al. Pharmacologic Blockade of 15-PGDH Protects Against Acute Renal Injury Induced by LPS in Mice [J]. Frontiers in physiology, 2020, 11: 138. |
| [26] | Sun M, Li J, Mao L, et al. p53 Deacetylation Alleviates Sepsis-Induced Acute Kidney Injury by Promoting Autophagy [J]. Frontiers in immunology, 2021, 12: 685523. |
| [27] | Wang Y, Wang X, Wang H, et al. PTEN protects kidney against acute kidney injury by alleviating apoptosis and promoting autophagy via regulating HIF1-α and mTOR through PI3K/Akt pathway [J]. Experimental cell research, 2021, 406(1): 112729. |
APA Style
Yang, P., Si, C. (2025). Analysis and Validation of Autophagy Related Genes in Septic Acute Kidney Injury. Biomedical Sciences, 11(2), 24-35. https://doi.org/10.11648/j.bs.20251102.11
ACS Style
Yang, P.; Si, C. Analysis and Validation of Autophagy Related Genes in Septic Acute Kidney Injury. Biomed. Sci. 2025, 11(2), 24-35. doi: 10.11648/j.bs.20251102.11
AMA Style
Yang P, Si C. Analysis and Validation of Autophagy Related Genes in Septic Acute Kidney Injury. Biomed Sci. 2025;11(2):24-35. doi: 10.11648/j.bs.20251102.11
@article{10.11648/j.bs.20251102.11,
author = {Pan Yang and Chen Si},
title = {Analysis and Validation of Autophagy Related Genes in Septic Acute Kidney Injury
},
journal = {Biomedical Sciences},
volume = {11},
number = {2},
pages = {24-35},
doi = {10.11648/j.bs.20251102.11},
url = {https://doi.org/10.11648/j.bs.20251102.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.bs.20251102.11},
abstract = {Background: The pathogenesis of acute kidney injury (AKI) in sepsis involves inflammatory response. Autophagy has been shown to regulate inflammatory response, but the role of autophagy-related genes (ARGs) in the regulation of inflammation in septic AKI requires further investigation. Methods: Initially, the dataset GSE57065 was downloaded and utilized in the R language to investigate differentially expressed autophagy-related genes (DEARGs) associated with septic shock. Subsequently, DEARGs enrichment analysis and protein-protein interactions (PPIs) were conducted. Hub genes were identified through PPIs, and their diagnostic value was evaluated using ROC analyses with the external dataset GSE65682. Additionally, the septic AKI animal model was established to validate hub genes through qRT-PCR. Finally, immune cell infiltration in the septic AKI and control group was analyzed. Furthermore, we examined the correlation between immune cell infiltration and the validated hub genes, and predicted the miRNA-mRNA network. Results: In the GSE57065 dataset, we have identified 22 differentially expressed genes (DEGs) primarily involved in autophagy. The receiver operating characteristic (ROC) analysis suggested that the top ten hub genes may have diagnostic value. In our animal experiment, qRT-PCR results demonstrated elevated expressions of TP53, MYC, FOXO1, CXCR4, and BCL-2, while PARP1 and PTEN exhibited reduced expressions in septic AKI. The CIBERSORT analysis revealed immune infiltration, and the validated hub genes were associated with immune cell infiltration in septic AKI. Lastly, we have discovered potential regulatory miRNAs through the miRNA-mRNA network. Conclusions: The potential diagnostic and therapeutic targets, namely TP53, MYC, FOXO1, CXCR4, BCL-2, PARP1, and PTEN, have been identified as potentially influential factors in the infiltration of immune cells in acute kidney injury associated with sepsis.},
year = {2025}
}
TY - JOUR T1 - Analysis and Validation of Autophagy Related Genes in Septic Acute Kidney Injury AU - Pan Yang AU - Chen Si Y1 - 2025/08/26 PY - 2025 N1 - https://doi.org/10.11648/j.bs.20251102.11 DO - 10.11648/j.bs.20251102.11 T2 - Biomedical Sciences JF - Biomedical Sciences JO - Biomedical Sciences SP - 24 EP - 35 PB - Science Publishing Group SN - 2575-3932 UR - https://doi.org/10.11648/j.bs.20251102.11 AB - Background: The pathogenesis of acute kidney injury (AKI) in sepsis involves inflammatory response. Autophagy has been shown to regulate inflammatory response, but the role of autophagy-related genes (ARGs) in the regulation of inflammation in septic AKI requires further investigation. Methods: Initially, the dataset GSE57065 was downloaded and utilized in the R language to investigate differentially expressed autophagy-related genes (DEARGs) associated with septic shock. Subsequently, DEARGs enrichment analysis and protein-protein interactions (PPIs) were conducted. Hub genes were identified through PPIs, and their diagnostic value was evaluated using ROC analyses with the external dataset GSE65682. Additionally, the septic AKI animal model was established to validate hub genes through qRT-PCR. Finally, immune cell infiltration in the septic AKI and control group was analyzed. Furthermore, we examined the correlation between immune cell infiltration and the validated hub genes, and predicted the miRNA-mRNA network. Results: In the GSE57065 dataset, we have identified 22 differentially expressed genes (DEGs) primarily involved in autophagy. The receiver operating characteristic (ROC) analysis suggested that the top ten hub genes may have diagnostic value. In our animal experiment, qRT-PCR results demonstrated elevated expressions of TP53, MYC, FOXO1, CXCR4, and BCL-2, while PARP1 and PTEN exhibited reduced expressions in septic AKI. The CIBERSORT analysis revealed immune infiltration, and the validated hub genes were associated with immune cell infiltration in septic AKI. Lastly, we have discovered potential regulatory miRNAs through the miRNA-mRNA network. Conclusions: The potential diagnostic and therapeutic targets, namely TP53, MYC, FOXO1, CXCR4, BCL-2, PARP1, and PTEN, have been identified as potentially influential factors in the infiltration of immune cells in acute kidney injury associated with sepsis. VL - 11 IS - 2 ER -
Department of Pharmacy, Chongqing University Three Gorges Hospital, Chongqing, China
Department of Nephrology, Chongqing University Three Gorges Hospital, Chongqing, China
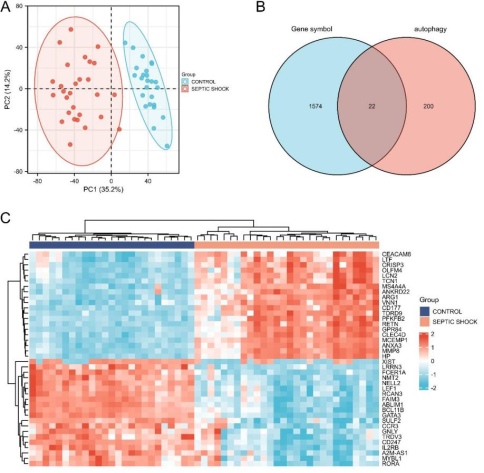
Figure 1. DEGs in septic shock group and control group. a Principal component analysis for GSE57065. b Venn diagram of intersecting genes between Human Autophagy Database and GSE57065. c Heatmap of the 20 up-regulated and down-regulated expressed genes in GSE57065.
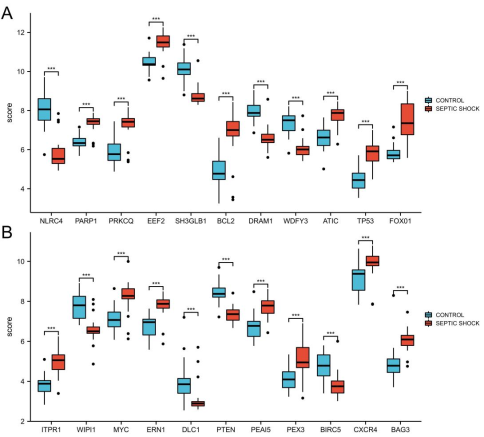
Figure 2. The boxplot of 22 DEARGs in septic shock group and control group. a The boxplot of top 11 DEARGs. b The boxplot of last 11 DEARGs. ***P<0.005.
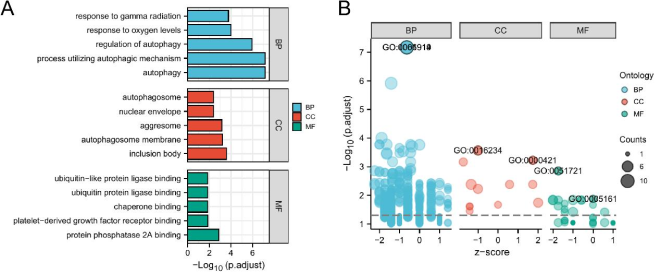
Figure 3. Gene Ontology (GO) enrichment analysis for autophagy related DEGs. a Barplot; b Dotplot.
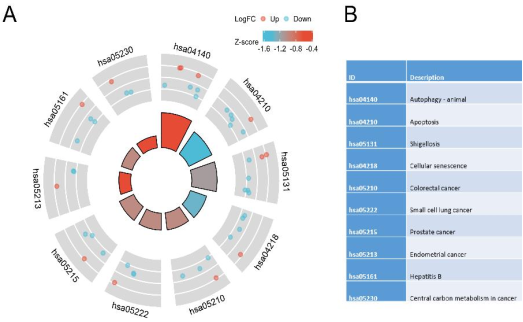
Figure 4. KEGG analysis of DEARGs.
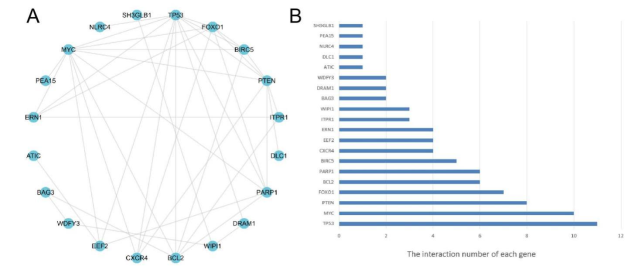
Figure 5. The PPI of the 22 DEARGs. a The PPI of the 22 DEARGs. b The interaction number of each DEARG.
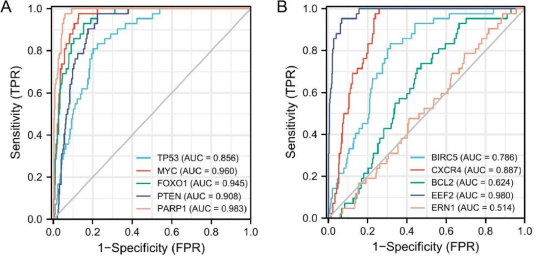
Figure 6. The ROCs of the hub genes. (A) TP53, MYC, FOXO1, PTEN, PARP1, (B) BIRC5, CXCR4, BCL2, EEF2, ERN1.
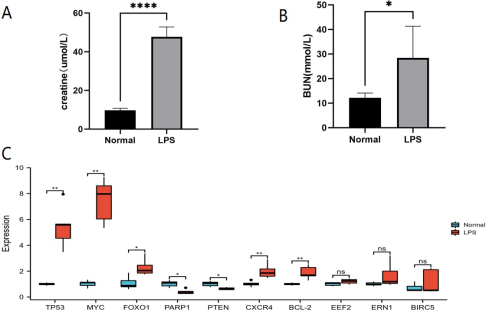
Figure 7. The validation of DEARGs in the animal experiment. (A) BUN and creatinine levels in mouse normal and LPS groups. (B) qRT-PCR analysis of the hub genes.
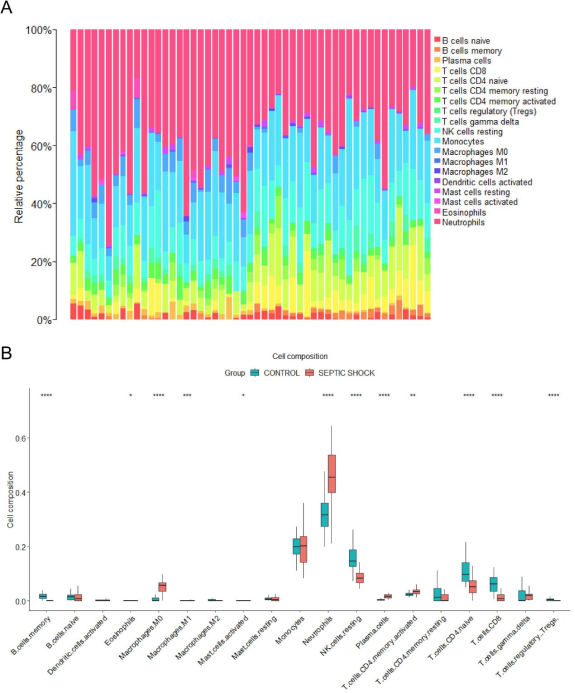
Figure 8. Analysis results of 22 kinds of immune cell infiltration. (A) Distribution of 22 kinds of immune cells in septic shock group and control group. (B) The heat map showed differences in immune cell infiltration.
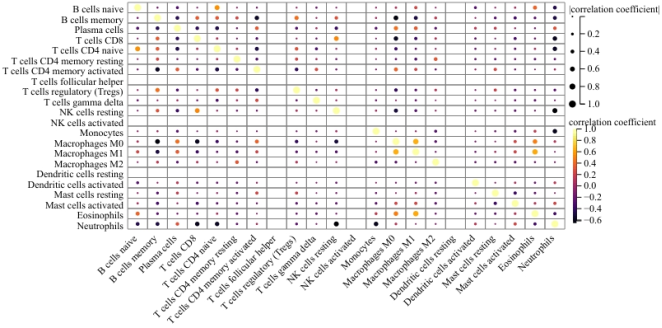
Figure 9. The heat map of immune cells. Blue and yellow indicated a negative and positive correlation between 22 immune cells, respectively. Dot size indicated closer ties.
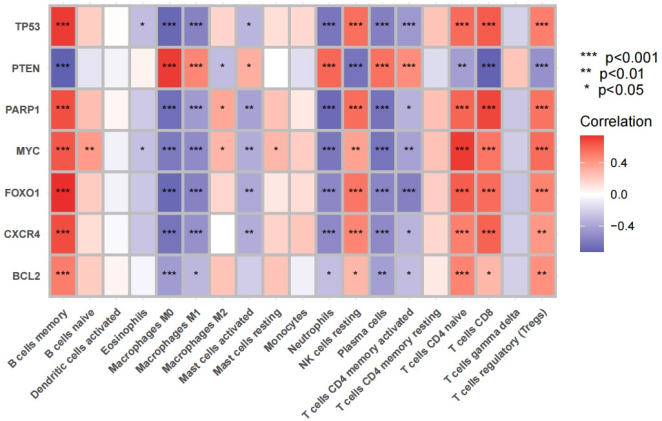
Figure 10. Correlation between TP53, PTEN, PARP1, MYC, FOXO1, CXCR4, BCL2 and immune cells.
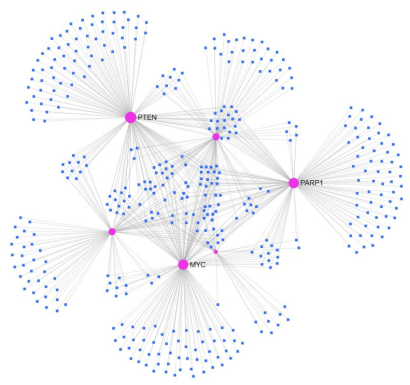
Figure 11. miRNA-hub genes network. Red represented hub genes, and blue represented miRNAs.
Information

